
Professor Kees Weijer FRSE
Chair of Developmental Physiology
Molecular Cell and Developmental Biology, School of Life Sciences

Contact
Research
Cell movement plays an essential role in the embryonic development of multicellular organisms, especially during gastrulation, formation of the nervous system and organogenesis. When not properly controlled it results in severe congenital defects and disease in later life. Cell movement is often guided by gradients of diffusible attractive and repulsive signalling molecules (chemotaxis). Important open questions are how do cells after detection of gradients of chemo-attractants/repellents translate this information in polarised organisation of the actin myosin cytoskeleton to result in directed movement up or down these signalling gradients?. A further key question is how chemotactic signalling and cell movement interact to create new tissues? We study these questions using a variety of multidisciplinary approaches in two model organisms, the genetically tractable social amoebae Dictyostelium discoideum and during gastrulation in the chick embryo.
Dictyostelium cells show very efficient homologies recombination, allowing the fast generation of targeted gene disruptions and replacements. We use these techniques in combination with high resolution SILAC based proteomics, TIRF and confocal imaging, dedicated microfluidic devices and detailed mathematical modelling to to study the molecular basis of serpentine receptor mediated chemotactic signal transduction to and organisation of the actin-myosin cytoskeleton (fig. 1). Dictyostelium cells do not only detect and move in response to the chemo-attractant cAMP, but they can also amplify and pass on cAMP signals to neighbouring cells, a process that results in the generation of propagating waves of cAMP (fig. 1). We study how these interactions between cAMP wave propagation and cell movement result in emergent behaviours and morphogenesis at the tissue level using again molecular genetics, advanced imaging and detailed mathematical modelling.
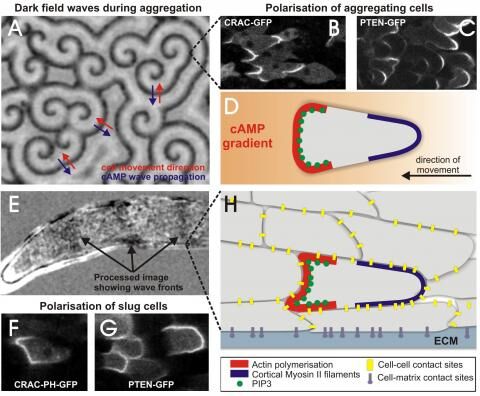
Figure 1: Aggregation of Dictyostelium cells is coordinated by propagating waves of cAMP, which can be visualised through their associated dark field waves. These darkfield waves reflect bands of moving and non-moving cells. The waves propagate from the centres outward (blue arrows in A) and direct the inward motion of the cells towards the aggregation centre (red arrows). Cells in a cAMP wave polarise, they produce PIP3 in the front (B, D), which is stabilised by PTEN localising to the back of the cells (C). Cell movement in slugs is also organised by waves propagating from the tip to the slug posterior and in slugs (E) Slug cells also polarise by making PIP3 (F) in the front of the cells and localising PTEN to the back (G). Slug cells make cell-cell and cell-matrix contacts that are required for getting traction during movement (H).
To study cell movement during gastrulation in the chick embryo we use a combination of advanced in-vivo imaging methods and computational data analysis and modelling techniques. We have designed and build a lightsheet microscope that allows us to image all (>100.000) cells in the early chick embryo and to analyse their individual behaviours over extended periods of time (fig 2). These massive data sets allow us to understand how the behaviour of so many cells is coordinated and integrated to result in the large scale tissue flows and deformations that underlie the formation of the primitive streak. Furthermore we are characterising the movement of mesoderm cells after their ingression through the streak (fig 3). Our results show that their complex movements appears to be controlled by a sequential combination of positive and negative chemotactic responses to different FGF’s and VEGF’s and also involves other as yet uncharacterised chemo-attractants.

Figure 2: Light sheet microscopy to study tissue dynamics during chick gastrulation. A: schematic of lightsheet microscope showing the orthogonal illumination and imaging axes. B: upper image sequence showing dividing cell (red dot) at 10x, lower image sequence showing ingressing cell (red dot). C: Overview image of early chick embryo derived from 3000 optical sections take every 2 minutes. The arrows indicate the direction and magnitude of epiblast tissue flow as calculated by Particle Image Velocimetry (PIV). D: blow up of centre of vortex show in red box in C
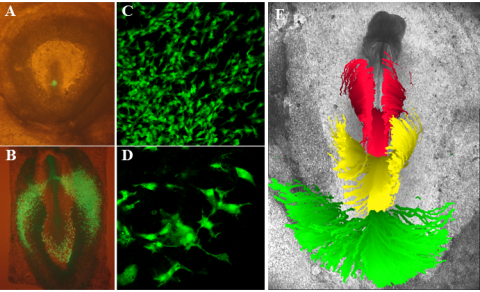
Figure 3: Migration of cells in mesoderm cells during the early stages of gastrulation. A: Image of a stage HH3 embryo directly after grafting a piece of middle streak tissue from a transgenic GFP expressing chick embryo of a similar stage in a homotypic position photograph of cell shape of GFP graft. B: Image of the same embryo after 20 hours incubation in culture. Most of the cells have migrated out of the streak where they will form the vasculature in the Area Opaca. The most anterior cells have stated to migrate back to participate in the formation of the embryonic vasculature. C: Higher magnification image of cells in the process of migrating out of the streak. It can be clearly seen that the cells are in close contact and highly polarised in the direction of migration. D: High magnification image of a few mesoderm cells at the edge of the cohort. Notice the very extensive cellular processes. E: Composite image of typical migration patterns observed of cell migrating from different antero-posterior positions in the streak during development from stage HH4 to stage HH7 (20 hours). Cells in the anterior streak migrate outward and then back in to form the somites (red tracks). Cells from the middle streak migrate out to form lateral plate mesoderm (yellow tracks) and cells located in the posterior streak migrate out in the extra-embryonic area to form the vasculature (green tracks).
Teaching
-
BS31004 - Biochemistry and Cell Biology
-
BS32005 - Developmental Biology
Awards
| Award | Year |
|---|---|
| Member of the European Molecular Biology Organisation | 2023 |
| Fellow of the Royal Society of Edinburgh | 2004 |
Stories
- Type
- Press release
Dundee scientist elected to elite scientific association
Professor Kees Weijer has been chosen to join the prestigious European Molecular Biology Organisation (EMBO), an organisation of leading researchers that promotes excellence in the life sciences in Europe and beyond.

- Type
- News
Further detail of early embryonic development uncovered
Latest research from the Universities of Dundee and Bristol has determined key details on the formation of large-scale functional structures during early embryonic development.

- Type
- News
Guillermo Serrano Najera awarded 2022 BSDB Beddington Medal
Guillermo Serrano Najera was awarded the British Society for Developmental Biology (BSDB) 2022 Beddington Medal at the recent annual meeting of the society. Guillermo received this awarded for his PhD work in Professor Kees Weijer on chick gastrulation.
